Original
Estimación del intervalo post-emersión de un cadáver hallado en un embalse en Granada (España)
Estimation of post-emersion interval of a corpse found in a reservoir in Granada (Spain)

Cuad Med Forense. 2011; 17(3):137-144
Fecha de recepción: 15.MAR.2011; Fecha de aceptación: 5.SEP.2011
RESUMEN
El período de actividad de los insectos es un parámetro que ha demostrado ser de gran utilidad a la hora de realizar aproximaciones a distintos fenómenos cronológicos relacionados con la descomposición cadavérica. Exponemos el caso de un cadáver hallado a finales de enero de 2011 en la orilla de un embalse en la provincia de Granada. Por la zona en la que se recuperaron los restos, la presencia de saponificación en la parte dorsal del tórax y la maceración de manos y pies, se sospechó inmediatamente de una sumersión de los restos y posterior emersión de los mismos. Tras recoger las evidencias entomológicas, se pudo determinar que consistían en larvas en diferentes fases de desarrollo de Calliphora vomitoria (Linnaeus, 1758), siendo las más antiguas recogidas en el cadáver las larvas en fase de desarrollo III migratoria. A partir del tratamiento estadístico de la información meteorológica recogida en dos estaciones agroclimáticas cercanas y del conocimiento del desarrollo y de la distribución de esta especie necrófaga, se pudo estimar que el tiempo mínimo que transcurrió desde que el cuerpo emergió del agua hasta que se encontró fue de 22 días. El díptero de la especie antes mencionada es compatible con la zona biogeográfica donde fue hallado el cadáver.
Palabras clave: ADD. Calliphora vomitoria. Entomología Forense. Intervalo postmortem. Cadáver sumergido.
ABSTRACT
Period of insect activity is a parameter that has been shown to be of great usefulness at the moment of making approximations to different chronological phenomena related to cadaveric decomposition. We expose the case of human remains found at the ending of January 2011 in the shore of a reservoir in the province of Granada. Because of the zone in which the remains were recovered, saponification in the dorsal area of the thorax, feet and hands maceration, submersion and a later emersion of the remains were immediately suspected. After collecting the entomological evidences, it was possible to determine that they were consisting of different developmental stages larvae of Calliphora vomitoria (Linnaeus, 1758), being the most aged ones recovered from the corpse corresponding to the larva III postfeeding developmental stage. From the statistical treatment of the meteorological data registered in two nearby agroclimatic stations and the knowledge of development and distribution of this necrophagous specie, it was possible to estimate that the minimum time elapsed from the moment when body emerged from the water until it was recovered was of 22 days. Diptera of the species previously mentioned is compatible with the biogeographic zone where the corpse was found.
Key words: ADD. Calliphora vomitoria. Forensic Entomology. Postmortem interval. Submerged corpse.
Introducción
La Entomología Forense es una disciplina científica que emplea los conocimientos sobre los ciclos vitales y la estructura de las poblaciones de artrópodos para esclarecer circunstancias relativas a casos que serán considerados en un tribunal de justicia. Dentro del vasto abanico de aplicaciones, una de las más conocidas es la estimación de la data de la muerte, denominada intervalo postmortem (IPM). Es importante tener en cuenta que el valor mínimo del IPM no siempre se corresponde con el periodo de actividad de los insectos, que es lo que realmente determina la Entomología Forense. El citado valor puede ser menor que el IPM cuando concurren circunstancias que puedan retrasar la colonización por parte de los hexápodos al cadáver o mayor en el caso de miasis1.
La principal herramienta con la que contamos en la determinación del IPM mínimo son las evidencias entomológicas. Los insectos son animales con una capacidad de regulación de la temperatura corporal muy limitada. Al no poder mantener una temperatura corporal constante, ésta dependerá en gran medida de la temperatura ambiente. El crecimiento de los insectos, por tanto, al ser un proceso dependiente de la temperatura corporal, dependerá en este caso directamente de la temperatura externa: a mayor temperatura, observaremos un desarrollo más rápido; si la temperatura decrece, la velocidad de crecimiento lo hará también2. Existen, además, dos umbrales de desarrollo que deben ser tenidos en consideración ya que el desarrollo se ve interrumpido por debajo (temperatura basal o umbral inferior de desarrollo) o por encima (umbral superior de desarrollo) de dichas temperaturas, pudiendo reanudarse cuando las condiciones ambientales vuelvan a ser favorables. De ahí que el conocimiento de la temperatura ambiental sea decisivo para unos correctos cálculos del periodo de actividad de los insectos.
Para poder conocer la edad de un estadio inmaduro de un insecto, recurriremos al conocimiento de las fases preimaginales de desarrollo. La mayor parte de las familias de dípteros de interés forense son ovíparas, es decir, la hembra grávida hace la puesta de huevos. De cada uno emerge una larva conocida como larva I. Cuando crece lo suficiente, pasa a larva II y luego a larva III. Posteriormente, la larva III se aleja de la fuente de alimento para pupar, conociéndose como prepupa o larva III migratoria. Cuando transcurre el tiempo necesario tras la pupación, emerge el adulto dejando tras de sí el pupario vacío3. Conocer estas fases y sus diferencias morfológicas es tremendamente importante ya que, para pasar de una fase a otra, es necesario un cierto tiempo para cada temperatura. Para la estimación del intervalo postmortem mínimo se consideran las larvas de mayor edad siempre que no sean individuos aislados ya que no podemos olvidar la posibilidad de una eclosión precoz de algunos huevos4.
Una forma de facilitar los cálculos es unir en una misma expresión la temperatura ambiente y el tiempo de desarrollo. Con esa finalidad, se crearon los grados-día acumulados y los grados-hora acumulados (ADD y ADH)5. Se pueden adoptar diferentes fórmulas para calcular los ADDs6, aunque una forma bastante útil en la que se da una compensación tanto por exceso como por defecto consiste en hacer el producto de la media de la temperatura diaria por la unidad de tiempo (1 día) y restarle la temperatura basal o umbral inferior de desarrollo como factor de corrección. Tiene un carácter más bien teórico ya que es una aproximación matemática a un modelo de crecimiento preseleccionado7. Para estimar cuándo comenzaron los insectos su desarrollo en el cadáver, calculamos los ADDs a partir de la suma acumulada de las unidades de energía expresadas en grados-día (DDs) que son requeridas por el espécimen considerado más antiguo que fue recogido del cadáver. La estimación se realiza de forma retrospectiva en el tiempo, desde el día del levantamiento hasta que se alcanzan los ADDs denominados «de referencia», obtenidos de datos publicados sobre la cría del espécimen en el laboratorio y en un régimen de temperaturas constantes.
Se define la sumersión como el conjunto de trastornos fisiopatológicos producidos por la entrada de un medio líquido en las vías respiratorias8. Aunque no es el objetivo de este artículo profundizar en la causa de la muerte, simplemente destacaremos que al diagnóstico de sumersión vital se puede llegar mediante el hallazgo durante la práctica de la autopsia de ciertos signos macroscópicos y/o diferentes tipos de pruebas complementarias9,10.
Los fenómenos destructores del cadáver aparecen de forma secuencial tanto en cadáveres sobre el terreno como en sumergidos mientras no aparezcan otros factores que retrasen o interrumpan la marcha de la degradación orgánica1. No obstante, las fases de la putrefacción varían en función de la disposición del cadáver: al aire libre respecto a los cadáveres sumergidos o enterrados, en agua dulce respecto al agua salada… Suelen reconocerse cinco fases en la descomposición cadavérica a la intemperie: fresca, colorativa o cromática, enfisematosa, colicuativa y de reducción esquelética1. Entre otras características, cada una de ellas presenta eventos de transición (mancha verde, enfisema putrefactivo y marcaje de la red venosa superficial, colgajos epidérmicos y formación de serosidad con escape de gases, formación de putrílago y exposición de los restos óseos). La cronología en la que se instauran cada una de las etapas, lejos de ser constante, muestra una gran variabilidad basada en las vestiduras e influencias constitucionales, patológicas y ambientales.
Cuando el cuerpo se encuentra sumergido, las variaciones constitucionales y patológicas que se consideran en la descomposición sobre el terreno se mantienen. Las ambientales, sin embargo, se modifican por las características especiales del medio líquido. En este sentido, se considerará a la hora de evaluar la cronología de la descomposición la temperatura, salinidad, pH, depósito de materia orgánica (eutrofización), tamaño de partículas, turbulencia de aguas y estación11.
Los objetivos del estudio son la estimación del periodo mínimo de colonización en este caso y la demostración de que, aún en el caso de barreras físicas evidentes, se puede conseguir una aproximación a la data de la muerte basada en evidencias entomológicas.
Material y métodos
Descripción del caso
En enero de 2011 se encontró un cadáver en avanzado estado de descomposición en la orilla del embalse de Canales (Granada). Presentaba mancha verde generalizada, red vascular muy definida, zonas de inicio de adipocera, maceración de piel y faneras y entomofauna sarcosaprófaga (Figura 1). Debido a los procesos putrefactivos, la identificación positiva a partir de los rasgos faciales era imposible, pero se sospechó que se trataba de un varón que desapareció de su domicilio a finales de noviembre del año anterior ya que, según informó la Policía, la zona era frecuentada por el desaparecido desde que inició los trámites de divorcio. La temperatura en el lugar del levantamiento era de 5oC.
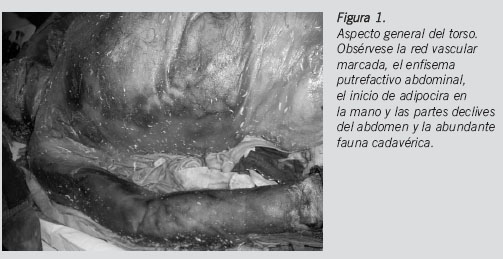
El lugar del levantamiento consistía en una cornisa de terreno que se levantaba por encima del pantano de Canales. El piso era de textura arenosa con algunas formaciones arbustivas de escasa frondosidad. Se describieron restos mucilaginosos de lo que pudieron ser algas que quedaron depositadas en el descenso de los niveles de llenado del embalse. Debido a la dificultad de acceso al lugar exacto donde se hallaba el cadáver, tuvieron que intervenir efectivos de Protección Civil para trasladar el cuerpo en lancha hasta un lugar seguro donde se continuaron las diligencias oportunas.
El caso que presentamos es paradigmático ya que se sospecha una inmersión del cuerpo previa a la colonización por parte de dípteros necrófagos y, por tanto, un retraso considerable entre el fallecimiento y la llegada de los primeros insectos de interés forense. A partir de los datos meteorológicos y el conocimiento de la secuencia temporal en el desarrollo larvario de una especie de díptero necrófago hallada sobre los restos humanos, se ha podido determinar el intervalo mínimo que transcurrió desde que el cuerpo emergió del agua hasta su hallazgo.
El cuerpo fue trasladado al Instituto de Medicina Legal de Granada, donde se le practicó la autopsia según la normativa europea12. La recogida de evidencias entomológicas durante la práctica de la autopsia y su interpretación en todas las fases de la investigación se realizó siguiendo las recomendaciones de la European Association for Forensic Entomology (EAFE)13. Las muestras fueron fotografiadas con testigo métrico y recogidas de la región cefálica por la médico forense encargada de la autopsia, se conservaron a una temperatura de 18oC y fueron trasportadas en recipiente isotérmico dos horas después por el responsable del análisis entomológico forense. Doce larvas vivas fueron criadas hasta la fase adulta en una incubadora Heraeus B-12, a una temperatura de 25oC, humedad relativa del 80% y alimentadas con hígado de cerdo ad libitum para realizar la identificación de la especie a partir de la fase adulta. El resto fueron sacrificadas sumergiéndolas en agua cercana a la ebullición durante 1 minuto y, posteriormente, conservadas en alcohol al 70% para la identificación de las larvas. De las larvas que fueron criadas, emergieron 9 adultos. Dos larvas de las que se remitieron muertas fueron sometidas a un aclaramiento en una disolución de hidróxido sódico al 10% m/v durante 24 horas. Posteriormente, se montaron en una preparación microscópica con DPX (VWR International®) para observar con mayor detalle las características morfoanatómicas. De los adultos, el muerto fue directamente conservado en alcohol al 70% y el otro sacrificado con acetato de etilo y posteriormente conservado con el mismo método que la muestra que se remitió muerta. La totalidad de las muestras fueron identificadas a nivel de especie14,15. No se recogieron muestras entomológicas durante el levantamiento del cadáver.
Datos ambientales
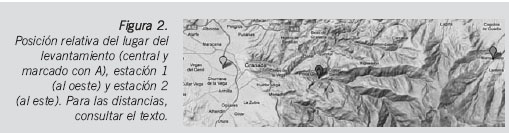
Los datos meteorológicos diarios (en el presente trabajo, tomamos las temperaturas diarias al no disponer de las horarias) fueron tomados de dos estaciones agroclimáticas distintas con el fin de evaluar cuál de ellas se ajustaba mejor a la temperatura del lugar del levantamiento16,17, ambas pertenecientes a la red de estaciones agroclimáticas del Instituto de Investigación Agraria y Pesquera de la Junta de Andalucía. La estación 1 (IFAPA de Camino del Purchil, coordenadas UTM X=443423.0 Y=4114357.0, altura sobre el nivel del mar: 630 metros) está situada a unos 13 kilómetros del lugar del levantamiento, la estación 2 (Jerez del Marquesado, coordenadas UTM, X=486806.0 Y=4116224.0, altura sobre el nivel del mar: 1212 metros) se halla a una distancia de 27 kilómetros del lugar del levantamiento (Figura 2). Para poder llevar a cabo la evaluación de ese ajuste de temperaturas, se colocó un data logger (ILOG S/INT-ESCORT®) en las proximidades del lugar del hallazgo del cuerpo debido a la imposibilidad de acceder al sitio exacto del levantamiento y se registraron las temperaturas máximas y mínimas durante cinco días, concretamente desde el 1 de febrero de 2011 hasta el día 5 del mismo mes. Los resultados fueron analizados con el paquete estadístico SPSS 15.0 y complementados con UNStat, aplicación desarrollada por la Universidad de Navarra.
Para diferenciar entre las larvas que todavía se están alimentando del cuerpo en descomposición y las que se están alejando ya de la fuente de alimentación para pupar, se diseccionan dorsalmente las larvas y se comprueba si existe un depósito de grasa subcuticular que impida la observación dorsal del tracto digestivo relleno de alimento. Ese depósito es observable en las larvas migratorias3. Además, la medición de la longitud del tracto digestivo relleno permite discernir entre ambas fases18. Mientras las larvas se alimenten, el contenido digestivo va aumentando su longitud de relleno mientras que, cuando entran en fase migratoria, existe un vaciado del contenido digestivo que implica un acortamiento de la longitud de tracto digestivo relleno.
Resultados
Fases del cadáver sumergido
Para el caso que nos ocupa, el cuerpo fue encontrado en asociación con un medio de aguas continentales lénticas (de flujo encerrado y lento). Para poder modelizar de forma adecuada la colonización de los restos cadavéricos, se adoptan cinco fases (sumergido fresco, flotación temprana, flotación en descomposición, flotación en descomposición avanzada y restos hundidos)19 y se desarrolla una gráfica basada en dicha referencia (Figura 3).
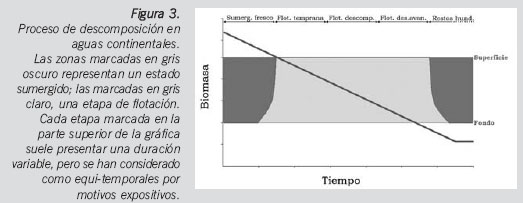
En el presente caso, el cálculo del intervalo post-emersión mínimo permite determinar cuándo finalizó la descomposición en medio acuático y cuándo comenzó la de tierra firme. No obstante, no podemos asegurar con precisión qué fenómenos cadavéricos corresponden a cada tipo de putrefacción. Según el esquema expuesto y atendiendo al lugar donde el cuerpo fue encontrado, la presencia de algas mucilaginosas adheridas al cuerpo y la incipiente saponificación, se puede elaborar la hipótesis de que el cuerpo permaneció sumergido hasta que entró en la fase de flotación temprana. Se plantean dos posibilidades: o bien el cuerpo abandonó la descomposición en agua en la fase de flotación temprana y el resto de fenómenos permitieron el desarrollo hasta una fase enfisematosa avanzada o bien se llegó en agua hasta la flotación en descomposición y el resto de fenómenos cadavéricos en tierra permitieron alcanzar un estado más avanzado de la fase enfisematosa en la que se encontraron los restos.
Análisis de los datos meteorológicos
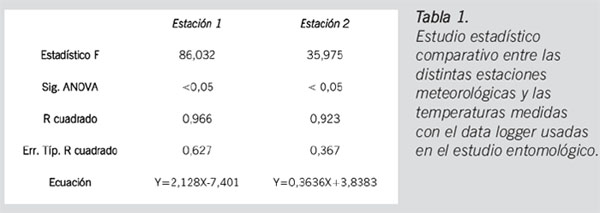
Lo primero que se debe considerar es si los datos registrados por las estaciones meteorológicas durante los días anteriores al hallazgo del cuerpo se ajustan a la realidad climatológica de dicho lugar. Para ello, como ya se indicó en el apartado anterior, se tomaron temperaturas durante 5 días en la zona del levantamiento y se procedió a realizar un análisis de regresión con las temperaturas para los mismos días en las estaciones. Para la elección de la estación que vamos a emplear hemos empleado el coeficiente de determinación (R2) más alto. Los resultados de dicho estudio estadístico se ofrecen en la Tabla 1. El valor de la R2 más elevado es de 0,966, es decir, del 96,6% de la variabilidad observada en las temperaturas del lugar de los hechos es posible inferir las temperaturas de la estación agroclimática del IFAPA de Camino del Purchil.
Análisis de las evidencias entomológicas
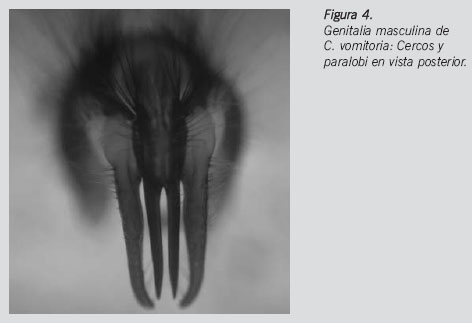
Se recogieron un total de tres tipos de evidencias entomológicas: uno constaba de dos adultos (uno vivo y otro muerto), otro de 35 larvas muertas en alcohol al 70% y el último de 27 larvas vivas con sustrato alimenticio. De los 9 adultos que emergieron a partir de las 12 larvas criadas en condiciones controladas, se identificaron 3 machos y 6 hembras. Todos los ejemplares pertenecían a la familia Calliphoridae. La identificación de los machos adultos fue confirmada mediante el estudio de las genitalias (Figura 4). La totalidad de las muestras pertenecen a la especie Calliphora vomitoria (Linnaeus, 1758) (Diptera, Calliphoridae).
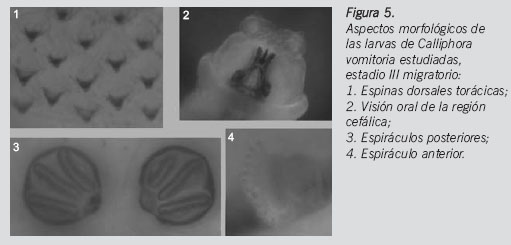
Sobre el frasco que contenía larvas muertas, 22 se encontraban en fase III migratoria, 10 en larva III alimenticia y 3 en larva II. De las 15 larvas restantes en el frasco de larvas remitidas vivas que fueron sacrificadas, 6 eran larvas III migratorias y 9 larvas III alimenticias (Figura 5). La identificación de las larvas es confirmada por el estudio de los adultos que emergieron de la cría controlada, ayudándonos en el caso de los machos del estudio morfológico de las genitalias.
Data del intervalo post-emersión
Basándonos en experiencias previas en la cría bajo condiciones controladas de especímenes de C. vomitoria, utilizaremos los datos sobre desarrollo publicados por Greenberg y Tantawi20 considerando una temperatura basal de 3oC20,21, ya que son los que más se ajustan a nuestros resultados y se pueden consultar en una búsqueda bibliográfica (Tabla 2).

Las larvas de mayor edad halladas en el cadáver pertenecen a la fase III migratoria. Para esa fase, consideramos un ADD-B3 de referencia (ADD con temperatura basal de 3oC) de 181,4520,21. Marchenko21 recomienda que las larvas se críen en condiciones controladas hasta alcanzar la fase de pupa o el adulto. A partir del tiempo que han necesitado las larvas en la cámara de cría para alcanzar cualquiera de las dos fases de desarrollo anteriores, se calculan entonces los ADDs a esa fase (pupa o adulto) en cría controlada y el resultado se resta a los ADDs de pupa o adulto que se tienen en las tablas de referencia. Nosotros sólo hemos empleado la cría controlada para confirmar la identidad de las larvas ya que, al trabajar con datos bibliográficos, la variabilidad intraespecífica propia de las adaptaciones a la climatología local es un factor de error que ha de tenerse en cuenta e imposibilita una mayor precisión, precisión que es vital para operar con ADDs y realizar la sustracción.
Para el cálculo de la temperatura diaria en el lugar del levantamiento, emplearemos la ecuación de regresión obtenida por el ajuste de temperaturas entre la zona del embalse de Canales y la estación 1: Y=2,128X-7,401, siendo Y la temperatura que consideraremos en el lugar de los hechos y X la temperatura registrada en la estación 1 (Tabla 3).
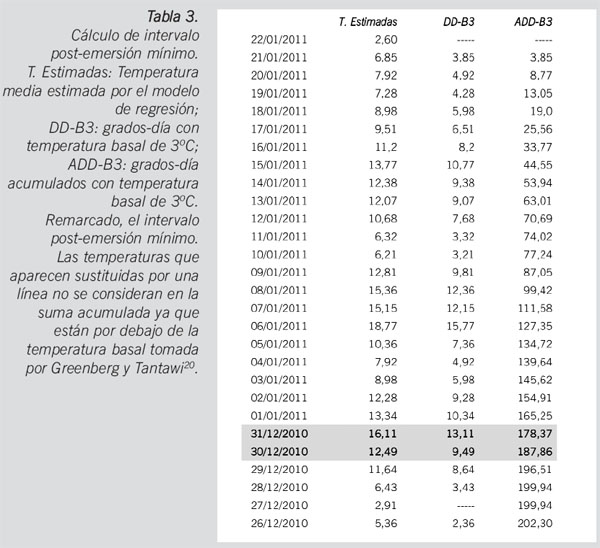
A tenor de lo expuesto anteriormente, el cadáver podría haber estado expuesto a la colonización del díptero citado durante 22 días, anteriores al momento en el que fue encontrado. El periodo postmortem mínimo calculado es inferior al periodo postmortem real ya que, por un lado, hemos considerado una temperatura de desarrollo de 12,5oC frente a una temperatura media del lugar del levantamiento de 10,81oC. Asimismo, solamente hemos considerado la llegada a larva III migratoria sin sustraer el tiempo de cría en condiciones controladas para la llegada a pupa, de modo que el tiempo de desarrollo considerado es inferior al real.
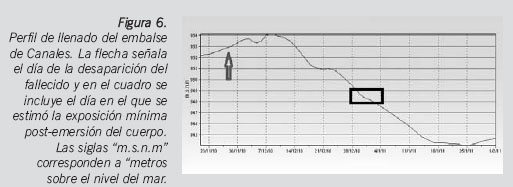
Con ayuda de un altímetro, se determinó que la altura sobre el nivel del mar de la región del embalse de Canales donde se halló el cuerpo era de 948,8 metros. Se hizo una búsqueda en el Sistema Automático de Información Hidrológica (SAIH) de la cuenca hidrográfica del Guadalquivir y se descargó un registro que incluía las fechas de desaparición de la persona a la que se le atribuía la identidad y la del hallazgo del cuerpo. Como se puede comprobar, el intervalo post-emersión mínimo que se calculó concuerda perfectamente con un descenso de los niveles de llenado del embalse y con la altura sobre el nivel del mar en esas fechas (Figura 6).
Discusión
Calliphora vomitoria es una especie holártica, de comportamiento algo menos heliófilo que especies del mismo género, termofóbica y cuyo máximo de abundancia en un gradiente altitudinal se encuentra en los 1.600-1.800 metros sobre el nivel del mar22. Debido precisamente a esa preferencia por los ambientes fríos, no es de extrañar que los picos de abundancia estacional se concentren, esencialmente, en otoño23.
Presenta un marcado carácter asinantrópico, entendiendo el concepto «sinantropía» como el grado de asociación entre una especie animal concreta y los asentamientos humanos24. Aunque se han descrito pocos casos sobre la aparición de C. vomitoria en cadáveres humanos25-28, puede llegar a ser la especie predominante en cuerpos hallados en sitios muy alejados de los asentamientos humanos, lo cual es compatible con el lugar del hallazgo del cuerpo. De hecho, se pueden establecer bastantes paralelismos entre el caso descrito por Wyss et al25 y el nuestro: en ambos está implicada la especie C. vomitoria, existe una barrera física para la puesta y desarrollo de los insectos inmaduros que implica el retraso en la oviposición por parte de la especie primaria, la estrategia utilizada es el estudio de los ADDs en lugar de los ADH (IPM mínimo de 6 días en el caso de Wyss) y se emplea la misma temperatura basal (3o C). Por otra parte, hay que tener en cuenta que también existen diferencias: en el estudio de Wyss se emplean tablas de crecimiento propias que no figuran en la publicación, hay una colonización por parte de Calliphora vicina Robineau-Desvoidy, 1830 posterior a la de C. vomitoria y se lleva a cabo la recomendación mencionada por Marchenko21 de considerar el tiempo de crecimiento controlado en laboratorio y sustraer los ADDs/ADHs por el mecanismo antes explicado. En nuestro caso, hemos empleado tablas de crecimiento que se puedan encontrar tras una revisión bibliográfica20,21.
Como gran parte de los miembros de la familia Calliphoridae, tanto los adultos como las larvas tienen un comportamiento claramente necrófago y coprófago. Si atendemos a algunos estudios sucesionales publicados en la Península Ibérica29,30, es una especie que suele hallarse en las estaciones frías y a partir de la fase enfisematosa de la putrefacción, estableciendo una clara diferencia con C. vicina, que se puede encontrar ya en la fase fresca de la descomposición. Hemos de tener en cuenta que, en este caso, la colonización por parte de C. vomitoria pudo producirse con bastante proximidad a la emersión del cuerpo ya que la descomposición de éste venía dándose desde la inmersión en el embalse.
Como se puede inferir de todo lo explicado sobre el desarrollo de los insectos de interés forense, es imprescindible un conocimiento lo más exacto posible de las temperaturas en el lugar de levantamiento. La única referencia que podemos tener del historial de temperaturas de un lugar concreto procede de las estaciones meteorológicas más cercanas. No obstante, se debe tener en cuenta que la estación meteorológica más próxima no tiene por qué ser la que mejor refleje las condiciones ambientales17. Para evaluar el ajuste entre los registros de las estaciones meteorológicas y las recogidas durante unos días en el lugar del hallazgo del cuerpo se pueden usar diversas estrategias como por ejemplo una t de Student entre los diversos grupos de comparación (las estaciones)17 o el coeficiente R2 de la regresión lineal simple16.
Agradecimientos
Nuestro más sincero agradecimiento a los revisores del presente artículo por el tiempo y esfuerzo dedicados, cuyos comentarios y sugerencias han contribuido sustancialmente a la mejora de la calidad del mismo.
Bibliografía
- Goff ML. Early postmortem changes and stages of decomposition. En: Amendt J, Campobasso CP, Goff ML, Grassberger M (eds.). Current concepts in Forensic Entomology. 1 ed. Berlin: Editorial Springer; 2010. pp 1-24.
- De Réaumur RAF. Observation du thermometre faites a Paris pendant l´anne 1753, comparees avec cells qui ont ete faites sous la ligne a Isle de France, a Alger et en quelque-unes de nos isles de l´Amerique. 1 ed. París: Mémoires de l´Academie Royale des Sciences de l´Institut de France; 1753.
- Greenberg B. Flies as forensic indicators. J Med Entomol. 1991;28:565-77.
- Wells JD, King J. Incidence of precocious egg development in flies of forensic importance. Pan-Pac Entomol. 2001;77(4):235-9.
- Nabity PD, Higley LG, Heng-Moss TM. Effects of temperature on development of Phormia regina(Diptera, Calliphoridae) and use of development data in determining time intervals in forensic entomology. J Med Entomol. 2006;43:1276-86.
- VanLaerhoven SL. Blind validation of postmortem interval estimates using developmental rates of blow flies. Forensic Sci Int. 2008;180(2-3):76-80.
- Ikemoto T, Takai K. A new linearized formula for the law of total effective temperature and the evaluation of line fitting methods with both variables subject to error. Environ Entomol. 2000;29:671-82.
- Concheiro Carro L, Suárez Peñaranda JM: Asfixias mecánicas. En: Gisbert Calabuig Medicina Legal y Toxicología. 6 ed. Barcelona: Masson; 2005. pp 460-78.
- Horton BP, Boreham S, Hillier C. The development and application of a diatom-based quantitative reconstruction technique in forensic science. J Forensic Sci. 2006;51(3):643-50.
- Lorin de la Grandmaison G, Leterreux M, Lasseuquette K, Álvarez JC, de Mazancourt P, Durigon M. Study of the diagnostic value of iron in freshwater drowning. Forensic Sci Int. 2006;157(2-3):117-20.
- Merritt RW, Wallace JR. The role of aquatic insects in forensic investigators. En: Byrd JH, Castner JL (eds). Forensic Entomology: The utility of arthropods in legal investigations. 2 ed. Boca Raton: CRC Press; 2010. pp 271-319.
- Donat-Laporta E (trad.). Recomendación NoR (99) 3 del Consejo de Ministros de los Estados Miembros del Consejo de Europa sobre armonización metodológica de las autopsias médico-legales. Rev Esp Med Leg. 1999;23:86-7.
- Amendt J, Campobasso CP, Gaudry E, Reiter C, LeBlanc HN, Hall MJR. Best practice in forensic entomology – standards and guidelines. Int J Legal Med. 2007; 121:90-104.
- Rognes K. Blowflies (Diptera, Calliphoridae) of Fennoscandia and Denmark. Fauna Entomologica Scandinavica, 24. 1 ed. Leiden: E. J. Brill; 1991.
- Velásquez Y, Magaña C, Martínez-Sánchez A, Rojo S. Diptera of forensic importance in the Iberian Peninsula: larval indentification key. Med Vet Entomol. 2010;24:293-308.
- Archer MS. The effect of time after body discovery on the accuracy of retrospective weather station ambient temperature corrections in forensic entomology. J Forensic Sci. 2004;49(3):553-9.
- Dabbs GR. Caution! All data are not created equal: The hazards of using National Weather Service data for calculating accumulated degree days. Forensic Sci Int. 2010;202:49-52.
- Anderson GS. Minimum and maximum development rate of some forensically important Calliphoridae (Diptera). J Forensic Sci. 2000;45(4):824-32.
- Zimmerman KA, Wallace JR. The potential to determine a postmortem submersion interval based on algal/diatom diversity on decomposing mammalian carcasses in Brakish ponds in Delaware. J Forensic Sci. 2008;53(4):935-41.
- Greenberg B, Tantawi TI. Different developmental strategies in two boreal blowflies. J Med Entomol. 1993;30:481-4.
- Marchenko MI. Medicolegal relevance of cadaver entomofauna for the determination of the time of death. Forensic Sci Int. 2001;120:89-109.
- Baz A, Cifrián B, Díaz-Aranda LM, Martín-Vega D. The distribution of adult blow-flies (Diptera: Calliphoridae) along an altitudinal gradient in Central Spain. Ann Soc Entomol Fr. 2007;43(3):289-96.
- Martínez-Sánchez AI, Rojo S, Marcos-García MA. Annual and spacial activity of dung flies and carrion in a Mediterranean holm-oak pasture ecosystem. Med Vet Entomol.2000;14:56-63.
- Gabre RM, Abou Zied EM. Sarcosaprophagous flies in Suez province, Egypt – II Synanthropic and abundance degrees. Bull Ent Soc Egypt. 2003;80: 125-32.
- Wyss C, Cherix D, Michaud K, Romain N. Pontes de Calliphora vicinaRobineau-Desvoidy et de Calliphora vomitoria (Linné) sur un cadavre humain enseveli dans la neige. Revue internationale de Criminologie et de Police technique et scientifique. 2003;56(1):112-6.
- Benecke M. Six forensic entomology cases – description and commentary. J Forensic Sci. 1998;43(4):797-805.
- Schroeder H, Klotzbach H, Püschel K. Insects’ colonization of human corpses in warm and cold season. Leg Med (Tokyo). 2003;5(1):372-4.
- Turchetto M, Lafisca S, Constantini G. Postmortem interval (PMI) determined by study sarcophagous biocenoses: three cases from the province of Venice (Italy). Forensic Sci Int. 2001;177(1):28-31.
- Castillo Miralbes M. Estudio de la entomofauna asociada a cadáveres en el Alto Aragón (España). 1 ed. Zaragoza: Monografía de la Sociedad Entomológica Aragonesa; 2002; volumen 6.
- Arnaldos MI, Romera E, Presa JJ, Luna A, García MD. Studies on seasonal arthropod succession on carrion in the southeastern Iberian Peninsula. Int J Legal Med. 2004;118:197-205.
Dirección para correspondencia:
A. González Medina
Instituto de Medicina Legal
Avenida de Madrid s/n,
18012 Granada.
E-mail: agm@ugr.es